Our thinking in this
chapter makes a significant shift away from received opinion, and into
territory that will be unfamiliar to most readers. Some of its
implications will easily be missed on first reading. It is a
distillation of several different sources - all intellectually
challenging - but the power and scope of this new way of understanding
make it well worth the effort it will take.
One of the shifts I am making is an
important re-application of the principle of natural selection. We are
not concerned here with the evolution of species (where, as I have
argued elsewhere
(1), the principle of
natural selection may have been seriously misapplied) but with the
evolution of behaviour in the here-and-now. I am arguing that natural
selection - the principle that declares that serious mistakes in the
conduct of life get automatically deleted from the hereditary line - is
something akin to a universal constraint on living processes. It
defines for every creature a behavioural range within which variations
can occur whilst life is able to continue. It is the discipline to
which all life is subject, not merely on pain of death, but of the
extinction of the germ-line (i.e. the whole series of potential
descendants). Though we do not know exactly how the living cell
organises itself so as to satisfy this constraint, it clearly manages
to do so by some means or another.
In this
chapter we
are looking at what a living creature does within the range
of
behaviour allowed it by the law of natural
selection. There is a repertoire of patterns of self-adjustment that
enable the creature in question to stay alive and eventually to
reproduce itself. We are seeking to understand that particular kind of
intelligence that we human beings have in common with the single-celled
organisms. This is an intelligence not to be looked down upon, since
it has evidently made possible the survival of primitive organisms
throughout the millennia. Also, if you accept (as I do) that the
theory of evolution entails the historical relatedness of all living
species then you should accept the consequence: this primitive
bio-intelligence has provided the fundamental pattern out of which the
entire kingdom of multi-cellular plants and animals has succeeded in
evolving.
Biology of Cognition
I want to
consider
the behaviour of a single-celled animal (such as Amoeba Proteus) from
the vantage point of cognitive biology, as it has been developed in the
work of Humberto Maturana and Francesco Varela in the late 1960s and
early 1970s. These researchers, one a neurophysiologist and the other
a biologist and mathematician, invented the concept of auto-poiesis,which
they hold to be the defining characteristic of the living creature. It
connotes the organised activity which creates the components
needed to constitute its own organisation. We should note that
this definition captures the closed-cycle nature of the living process.
This, argue Maturana and Varela, is what effectively demarcates the
boundary between the living and the non-living.
One unique
feature
of living protoplasm is the manufacture of large, chemically complex,
individually sculpted molecules based on long carbon chains of various
patterns of construction. These molecules are precisely organised in
relation to one another in space and time. They exist in a milieu
which also contains a whole variety of simpler chemical components
whose concentrations are maintained within a remarkably narrow range
(corresponding, I have been told, to the chemical composition of the
sea
in some primordial era). More impressive, perhaps, is the fact that
this precise and detailed regulation is happening in the face of myriad
comings and goings. The large molecules (starches, lipids, proteins
and nucleic acids) have relatively stable locations, subject to a
process of continuing re-modelling and maintenance, but the elemental
and simple ionic complexes (sodium, potassium, calcium, chloride,
bicarbonate etc) seem to be skeetering about haphazardly, except that
their comings and goings through the outer membrane of the organism are
regulated by a variety of mechanisms.
Our main
focus here
will be two-fold. We are looking within the structure of the organism
for the characteristic patterns of event which are the
precursors of emotional intelligence; and we are looking at the
creature's overall behaviour - its readiness to cope with
life-threatening situations, the apparent foresight with which it
interacts with objects in its surroundings, and its seeming dedication
to keeping its own life processes in balance. All of this evokes in us
the sense that the creature knows what it is doing. More
specifically, I am interested in the relationship between these two
levels - how the processes inside the organism's boundary contribute to
its survival as a whole.
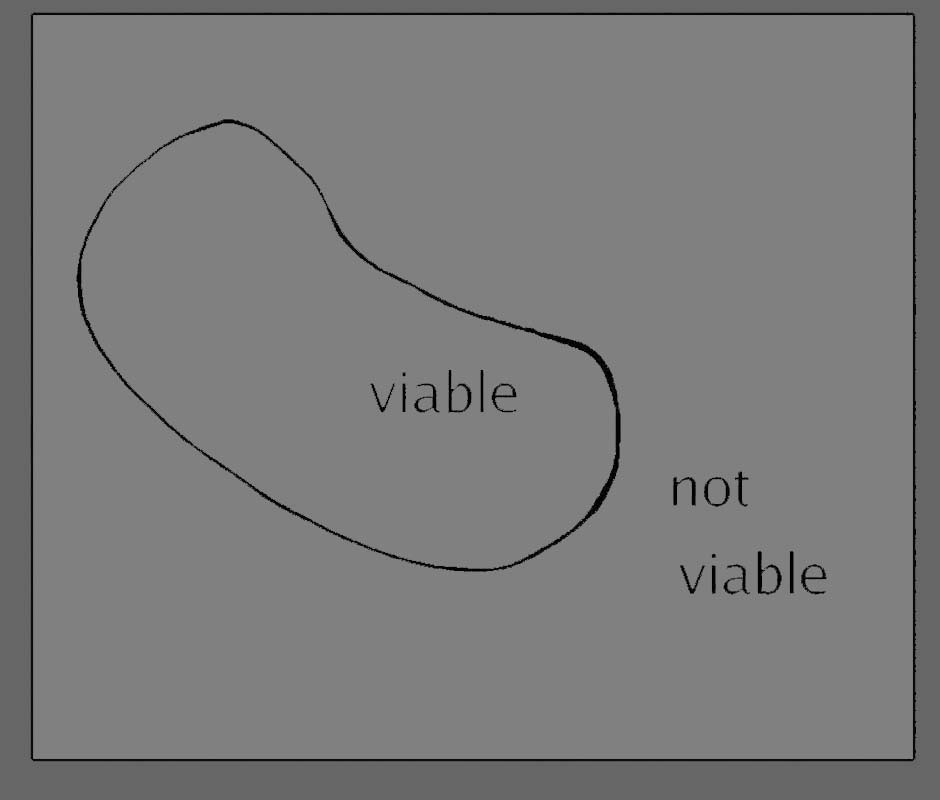
Figure
1
The account
which
follows has a number of diagrams attached and you need to be forewarned
that these are not quite what they seem. It is natural to interpret a
continuous, closed line such as we see in figure 1 as representing
something in physical space. This is not the case here, because we
are not interested in the physical boundary of the organism, as much as
in the boundary of its behaviour. These are therefore systems
diagrams (albeit very simple ones) which summarise and categorise an
enormous number of processes which happen within a given
period of time. The inside of the closed figure represents the "domain
of all possible internal states of the organism, compatible with its
being alive". The line itself represents the demarcation between the
states of the organism where it is still viable, and (on the outside of
the line) states that are not compatible with the continuation of life.
Each point within this boundary represents a determinate state of the
living organism. (Clearly I am not addressing any serious structural
detail in this diagram - a detailed behavioural map would require the
few square centimetres of our diagram to open up like one of those
Mandelbrot fractal pictures, in which more and more precise structure
emerges the closer we approach it.)
Any process
contributing to the life of the organism can be thought of as a
transition from one "state" to another "state". This event is
represented by a line passing from one point to another, the arrow
denoting a forward movement in time. (See figure 2.)
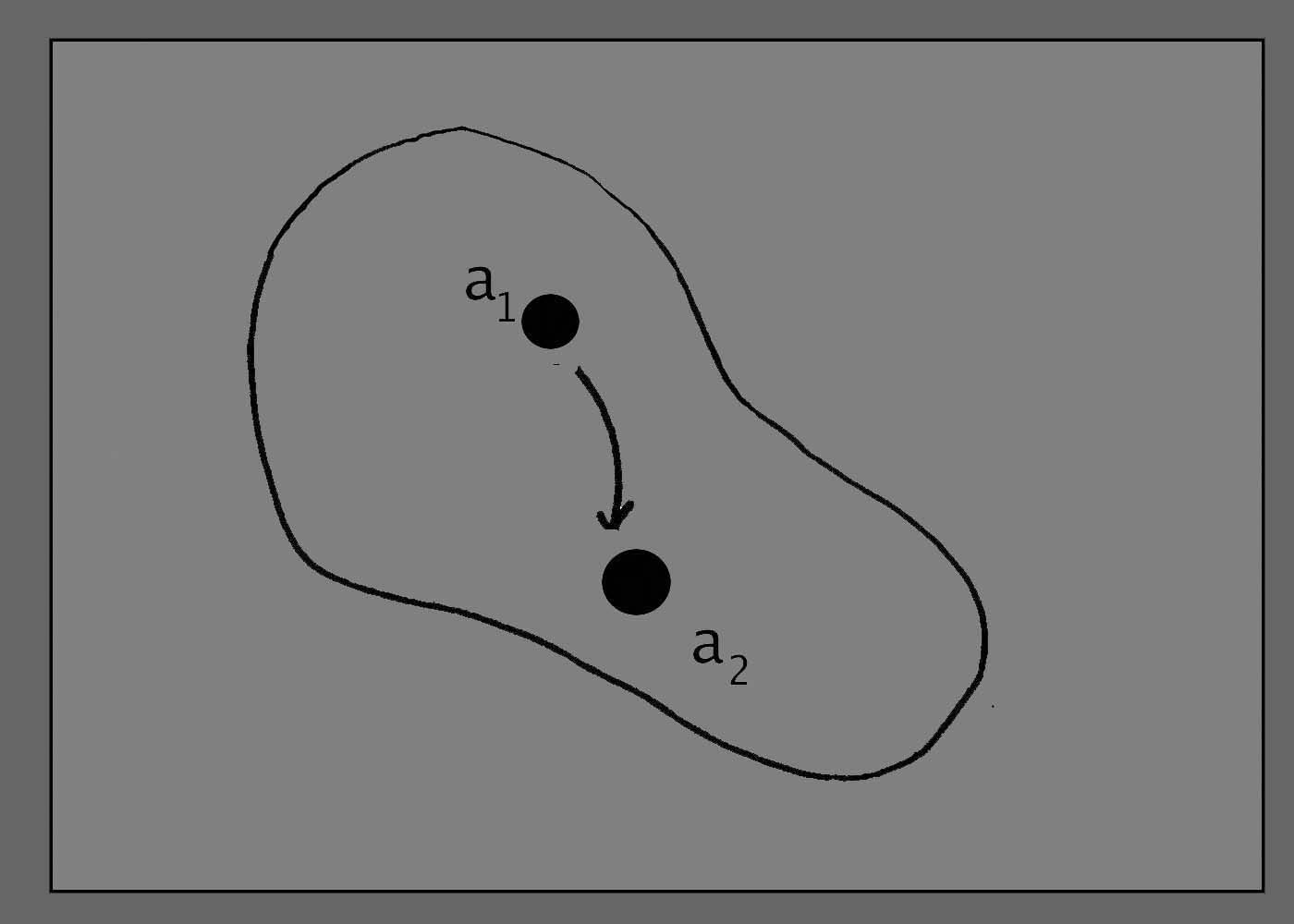
Figure
2
Since this is
a
lower-level process contributing to the higher-level configuration we
call "life", we need to recognize it as something more than a single
event. It is a recurrent event whose re-occurrence fulfils a
repeating function as one segment of a cyclical series of causes and
effects. There are two important points of contrast between this and
our common-sense notion of linear causation (which serves us so well in
the organisation of our practical life - in such activities as building
a house or designing a motor-car). They can best be demonstrated
visually, and so we shall represent the common-sense causation as a
straight-line series where one event directly influences the next in
the chain:-
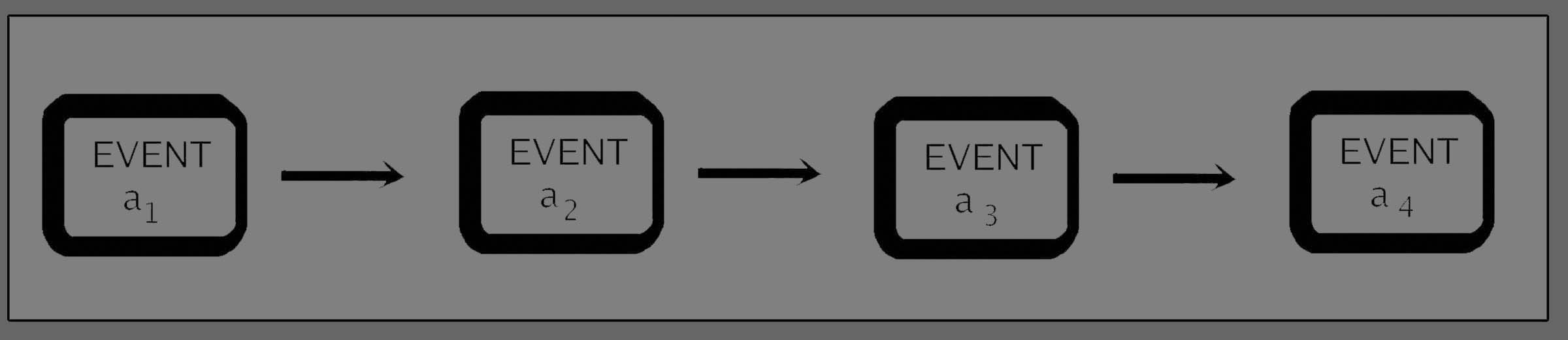
Figure
3
By contrast,
the
cyclical patterns characteristic of living processes have come to
arrange themselves in such a way that the outcome of any particular
physiological state is also a contributing factor in a chain of
causation which will lead to a recurrence of that same state
-
in a future cycle of events.
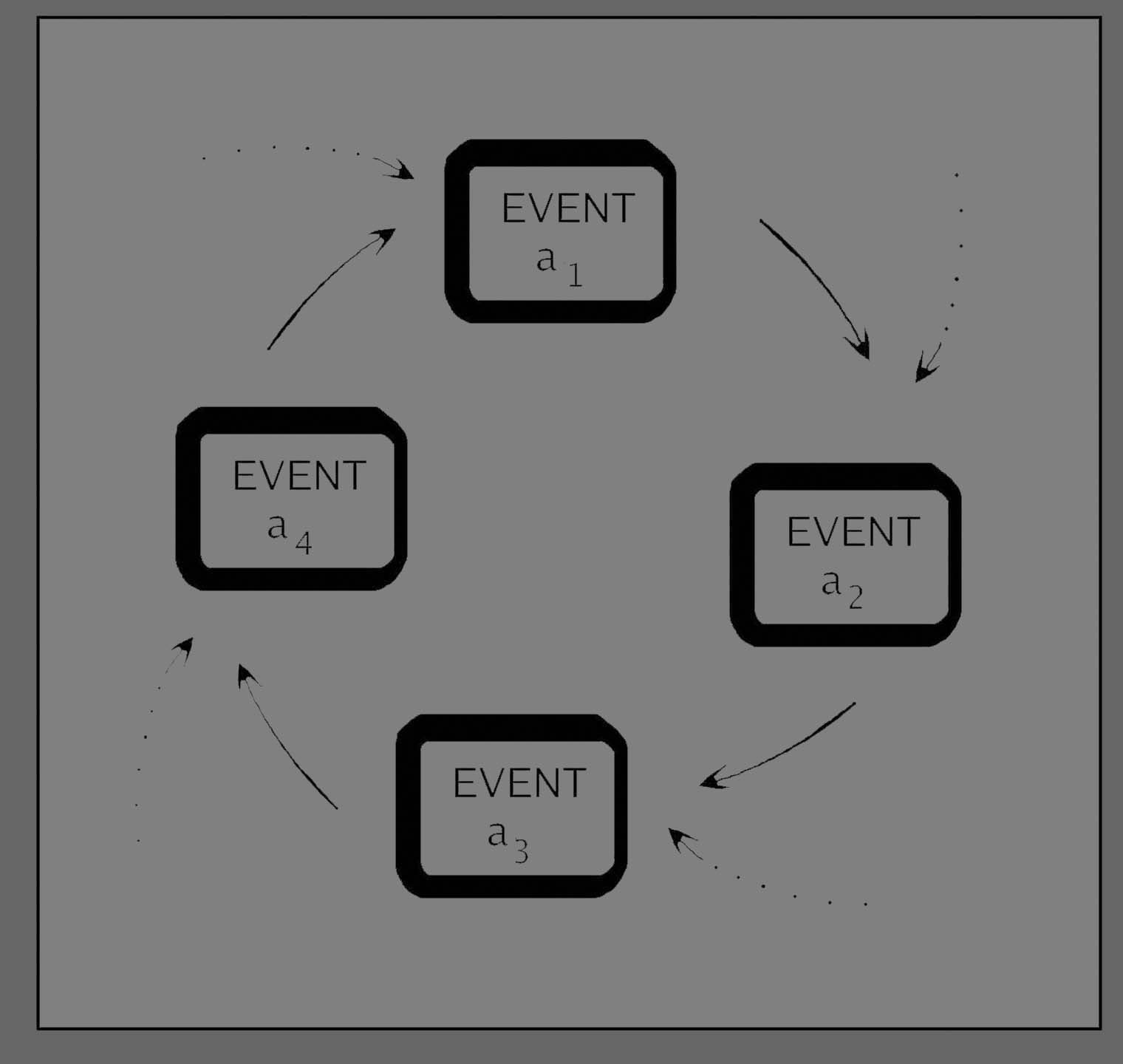
Figure
4
The second
contrast
is that these cyclical patterns often intersect with a great many other
similar ones - which means that any one event in the making, is subject
to a convergence of causes. This makes it impossible for us as
observers to say with certainty that event " a1"
actually caused event "a2" to occur; but we
may be reasonably sure that it entered into the causal ambience out of
which "a2" emerged. This is no different
from multi-factorial causation in other circumstances - such as a large
number of over-laden lorries crossing a bridge already weakened by
repeated attacks of frost, and the scheduled re-painting of the bridge
(which would have revealed dangerous fracture-lines in one of the
supporting arches) being delayed by the bad weather, leading to a major
collapse of the bridge.
Ultrastability
Certain
patterns of
circular causation have the additional property which systems engineers
call "ultrastability". An ultrastable pattern is one which repeats
itself in regular fashion, and is able to absorb the incidental impact
of a range of other events, so that its repetitions are maintained
within stable limits. All living organisms have this property and thus
continuously maintain their own internal balance under a range of
stresses. An important aspect of this property, which we might easily
overlook, is that the organism need not have any previous acquaintance
with - or understanding of - the specific nature of the disturbing
event in response to which it is having to restore its own equilibrium.
This differs from a typical computer-programmed activity, which
requires a complete specification of all relevant circumstances in
advance, with precise sets of instructions for how to deal with all
contingencies. The auto-poietic system need make no calculations in
respect of any world external to itself; it is enough that it measure
its own internal states and that it respond according to what it finds
within.
One example
of this
ultrastable cyclical pattern is that of DNA and RNA organising the
synthesis of protein which catalyses the process of sugar synthesis and
metabolism, which in turn provides the energy to maintain the whole
system which replicates DNA.
In his
classic
study of control and communications patterns applied to both living and
mechanical systems, "Decision and Control", Stafford Beer
argues that an essential component of the ultra-stable process is the
generation of some random variation at certain stages in the
process (but contained within the over-arching self-adjusting cycle).
This allows the system to find and then to amplify behavioural variants
which improve its stability through time. So even at this most basic
of levels of organisation, there is something like an exploratory or
creative function which can modify and improve the circular causal
organisation, without dismantling its essential continuity. This is
perhaps the crux of the circular practical logic underpinning the
living process - that it is the integrity of the cycle which
is the invariant in the system. The individual steps may be adjusted
and varied in all kinds of ways, on condition that the main cycle is
able to continue uninterrupted.
These basic
features need to be taken as read, when we look at "changes of state"
within the single-celled organism we are considering at the moment.
Thus the single transition a1 >> a2
as represented in figure 4 must be understood as one event within
an auto-poietic cycle. We, as observers, have singled it out for our
attention as if it began at (a1) and ended
at (a2), but it has taken its origin in
auto-poietic process and it is leading into further auto-poietic
process - if it did not, it would not belong in this diagram.
We now need
to make
a further distinction, between the range of states we could call
"physiologically comfortable" and those we would call "physiologically
stressed". The states lying within the zone of stress are deviations
from the optimum that are not so great as to be incompatible with life,
but great enough to be registered within the system in such a way that
they trigger further changes, operating in the direction of restoring
the equilibrium back to within the zone of comfort. (See figure 5,
below.) The change a1 >> a2
was a movement away from physiological comfort. a2
>> a3
was a restoration of comfort, but to another state (a3)
distinct from (a1). Of course this is not
meant to represent any specific biochemical pathway of events, but it
takes for granted that the living cell is already organised
to
make these serial and simultaneous biochemical adjustments, and that
its success in doing so has been the pre-condition for the million-year
long survival of this cell and its ancestral line.
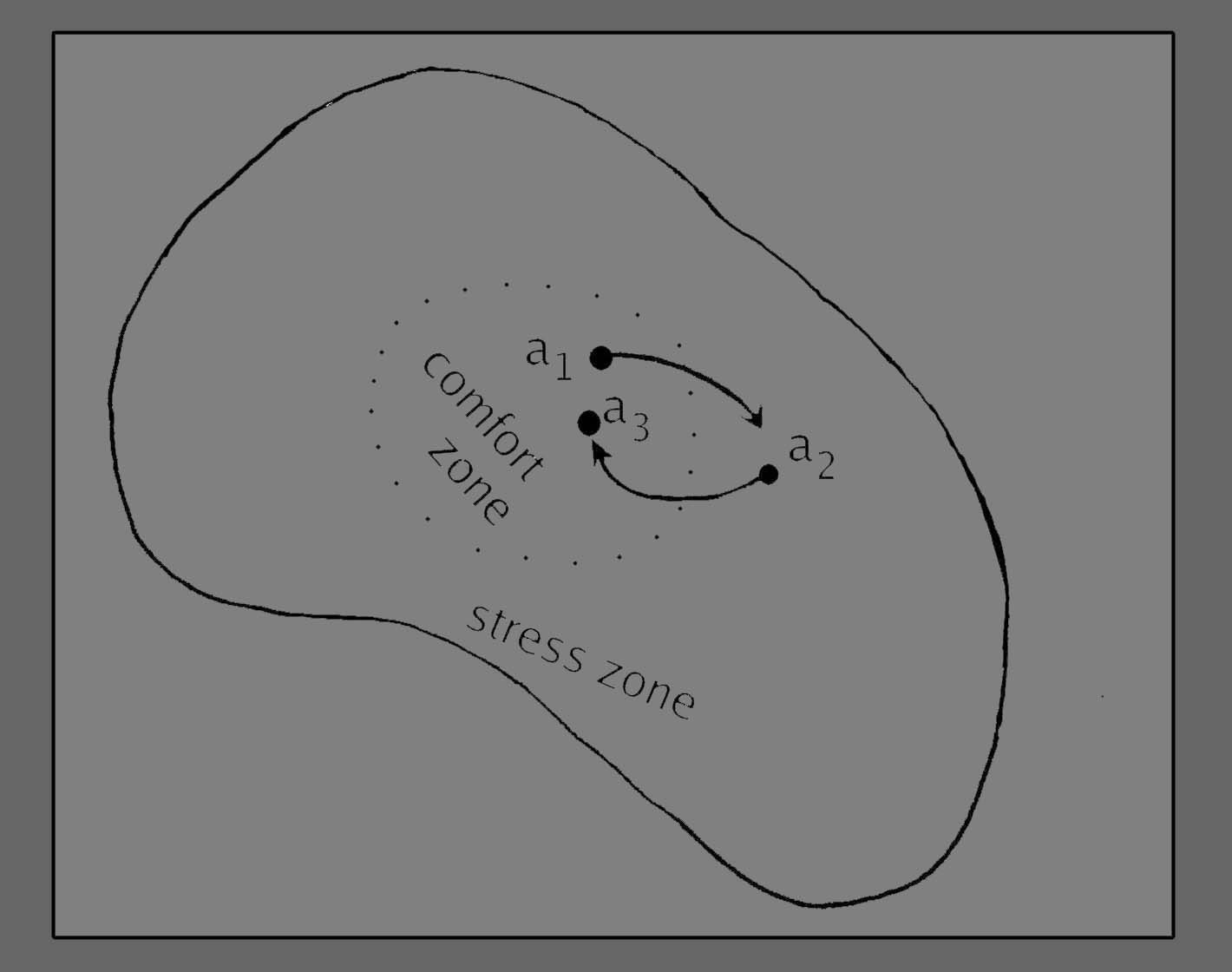
Figure
5
We know that
this auto-poietic process depends upon a myriad of interlocking,
cyclical biochemical pathways, a great many of which have been explored
by molecular biologists and biochemists over the past several decades.
Without the full complement of these biochemical processes the
auto-poietic cycle could not be maintained - but we do not need to
understand these processes in detail here.
This is in
exact
analogy to the correct notes needed to execute "I Can't Give You
Anything But Love!" and the convergence of strategies which achieved
the victory of Wellington's army at Waterloo - we recognize the overall
result but may be largely ignorant of the details.
The
continuous
monitoring of internal states, characteristic of the simplest of
organisms, is a close analogy with our own experience of feeling.
A physiological state which falls in the zone of stress corresponds to
a feeling of pain or discomfort. It signals that "something is wrong"
and that "something needs to be done". Looking back at our earlier
description, we could say that this feeling of stress enters into the
overall causal ambience so as to increase the probability that
subsequent changes in internal state will move the system closer to the
zone of physiological comfort.
In general,
the
longer, and the more seriously, our creature finds itself outside its
physiological comfort-zone, the more strongly its priorities become
diverted to the matter of restoring physiological comfort by whatever
means are available. We do not have to think of our little friend as
being aware of anything outside the domain of its own internal states.
It is enough that it registers its own states of physiological stress
and comfort, and has appropriate response pathways for maintaining
itself through the stresses and strains of its insecure existence.
Another
fundamental
concept we need to borrow from Maturana and Varela is that of structural
coupling. This is the automatic consequence of an auto-poietic
creature living within a milieu which contains needed raw materials and
potential threats and dangers. The creature is organised so that
outside influences are to a great extent buffered by the internally
driven ultrastable response patterns. In effect the organism is
resonating like an Aeolian harp in response to every impact from
outside. The pattern of this response is determined by the
organism's own structure; it is not like a direct imprint or impression
of the outside world. The effect through time is that the patterns of
event within the cell become spontaneously coupled with patterns of
event in the surrounding milieu with increasing precision - while their
detail remains determined moment by moment (in all important respects)
by the structure of the organism
Some details
of the
coupling will have developed over a span of many past generations; some
will have been acquired over this creature's own lifetime, but its
actual mechanism always operates in present time, as a consequence of
the organism's continuous ultrastable adjustment procedures.
The essential
point
about structural coupling is subtle, and depends upon our being clear
about the difference between our point of view as observers
of
the Amoeba in its natural medium, and the point of view of the Amoeba
itself. As observers we are able to interact with, and to keep track
of, both the amoeba and some of the events surrounding it. We are able
to recognize that events within the cell are moving in repeating
patterns which are effectively tracking the patterns of event in the
surrounding world. If we now shift in our imagination to try to
consider the creature's own point of view (within the framework of
understanding we have been developing here) we may be surprised to
realize that it has no need of any "information from
outside",
nor does it need to know that there is any such thing as an outside.
Its own auto-poietic activity (in the context of its structural
coupling with the surrounding medium) is giving it all the information
it needs, inscribed within the changing pattern of its own internal
states.
If we want an
analogy with something in our own life experience, this would be a
feeling about something which guides our action independently of
any need for conscious recognition of what we are doing(2).
We easily overlook such responses; our own are by definition unaware,
and when we read other people's responses to situations we
tend to attribute conscious perception to them just because our own
reading has been conscious. Consider the following, however: I am
approaching a bus-stop, walking in the direction of traffic so that I
see the people waiting and beginning to shift their positions in
response to a bus which I cannot see (since it is approaching from
behind me). I have several levels of response open to me. The
simplest is that I sense the approach of the bus without making any
conscious inferences at all. This is the response which is equivalent
to the level of structural coupling - the people at the bus-stop are
shifting around as if at the approach of a bus, and I take
in this ambience by a kind of osmosis, so that I too feel as if there
is a bus approaching. The conscious route to this awareness might be
that I notice people looking up and starting to move, and I look behind
me and see the bus for myself. My assumption now is probably that
"they saw the bus first", attributing a conscious perception to the
other people. The important insight for us here is that the people
would probably start to move anyway - regardless of whether they consciously
noticed
the bus or not. We are often so much in tune with surrounding events
that we come up with sensitive responses without any need for thought.
Returning to
our
single-celled organism, we see there is no need to attribute any
knowledge of the outside world at all - neither conscious nor
unconscious - since the relevant behaviours arise out of the sequences
of internal state, which are structurally coupled to patterns of events
on the outside. We do naturally think of the amoeba as responding to
objects in its environment when we watch its approach to food and
illumination, and when we notice the violent withdrawal of its
pseudopodia (the outward-flowing columns of protoplasm which extend
into limb-like processes during its normal movement in and around its
milieu) consequent upon physical impact or the presence of acid. But
it is also true at the level of the creature's own organisation that it
is simply "doing what comes naturally"; anything which impacts upon the
organism to induce a discernible change in its internal state will
provoke an auto-poietic response - which means a consequent change in
state that helps to maintain the system within its zone of
physiological comfort.
Rudiments of Language
Our sense
that the
creature "knows what it is doing" is all the more compelling when we
consider the dance which takes place between two amoebae in close
proximity. We cannot help but think of their very evident shifts and
adjustments as being "calls" and "responses" in relation to each
other. We need an extra conventional sign on our diagram (figure
6), which will stand for the shift in creature "A" as it impacts upon
the internal dynamics of creature "B". In the figure that depicts ‘serial interaction’ – which is Fig 7 – this shift is illustrated as a star, that is connected then to the projecting barbed line which passes from "A" to "B".
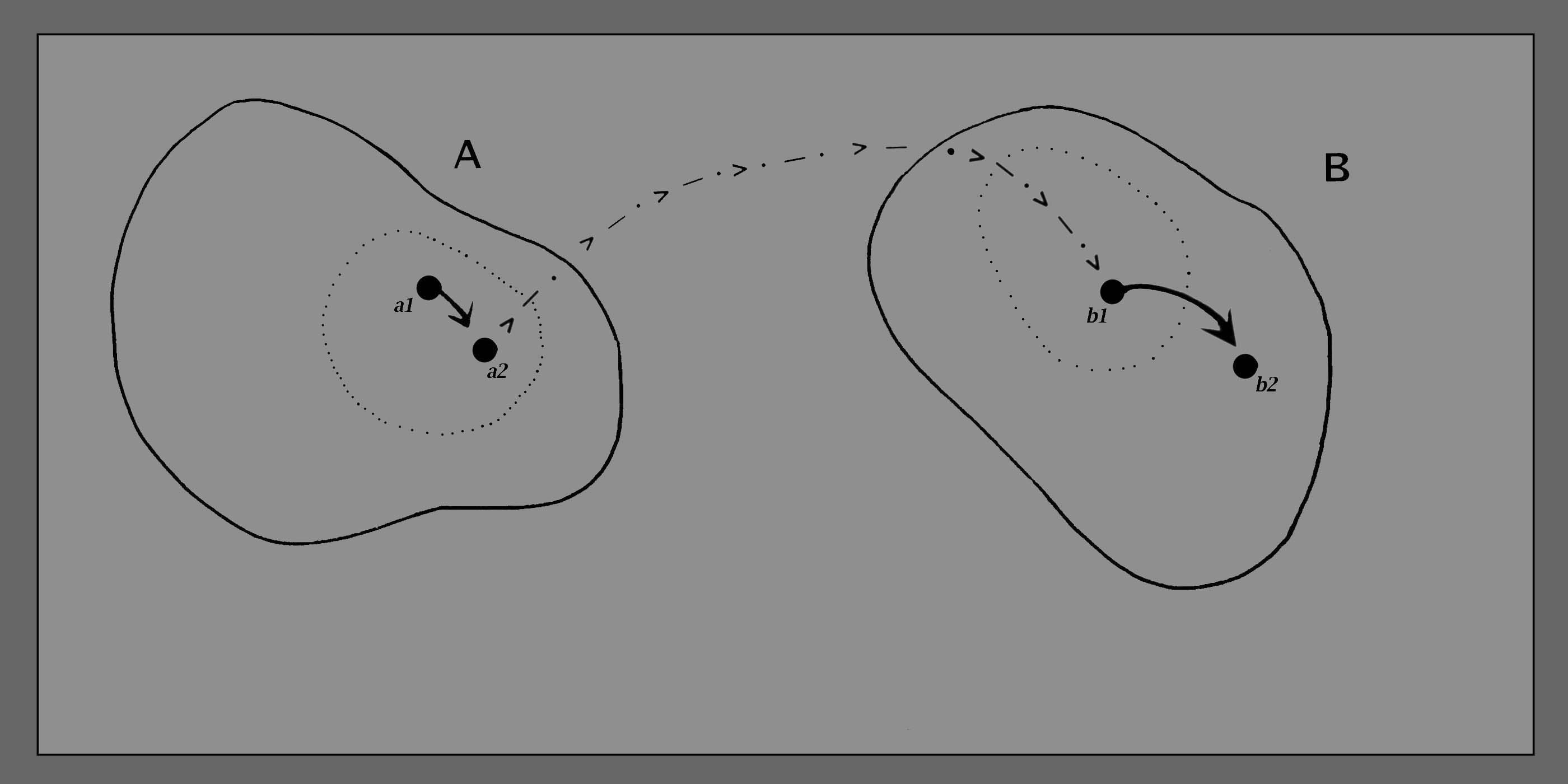
Figure
6
And in the
case
illustrated here it looks very much as if the change a1
>> a2 has triggered "B" into "becoming upset"
(b1 >> b2). This is
not necessarily the case. It is also
possible that the move from b1 to b2
could have arisen out of some important internal dynamic of "B" - or
indeed an interaction of several different factors. Therefore it would
not be correct to say that "a1 >> a2"
caused the subsequent shift "b1
>> b2". Rather we should say that "a1
>> a2" entered into the overall causal
ambience
out of which "b1 >> b2"
emerged.
At this level
of
description we are still not claiming any awareness on B's
part, of what has happened in A. We are simply saying that "B's" state
is significantly different from what it would have been, if a1
>> a2 had not happened. In Maturana and
Varela's
language, A is not imparting information to B, but is simply a source
of
"perturbations" (i.e. disturbances). In depicting this in diagram
form, we can make the assumption that every change in the
state
of "A" may have some impact on the subsequent change of state
which takes place in "B". ("No response" would thus be a special
case of the general situation "there will be a response of some
kind".)
We can now
move
forward to considering the case of serial interaction, in which each
shift in one organism acts as a possible trigger for a shift in the
other, which in turn acts as a possible trigger for the first.... and
so on until they either mate, wander off into separate parts of the
pond, or just continue to co-exist in a companionable way. This is
schematised in figure 7, below. There is no further mechanism required
to achieve this spontaneous process of continuous mutual adjustment
between the two creatures. They will learn, in effect, to take
advantage of whatever benefits might accrue from their mutual
proximity, and to make the best of any incidental disadantages. The
relationship will be more or less complex, depending upon the compexity
of the coupling between the sequence of states in the one creature, and
the sequence of states in the other. (The more "important" they are to
each other, the more intense and more precise will be the impacts of
the one upon the other.)
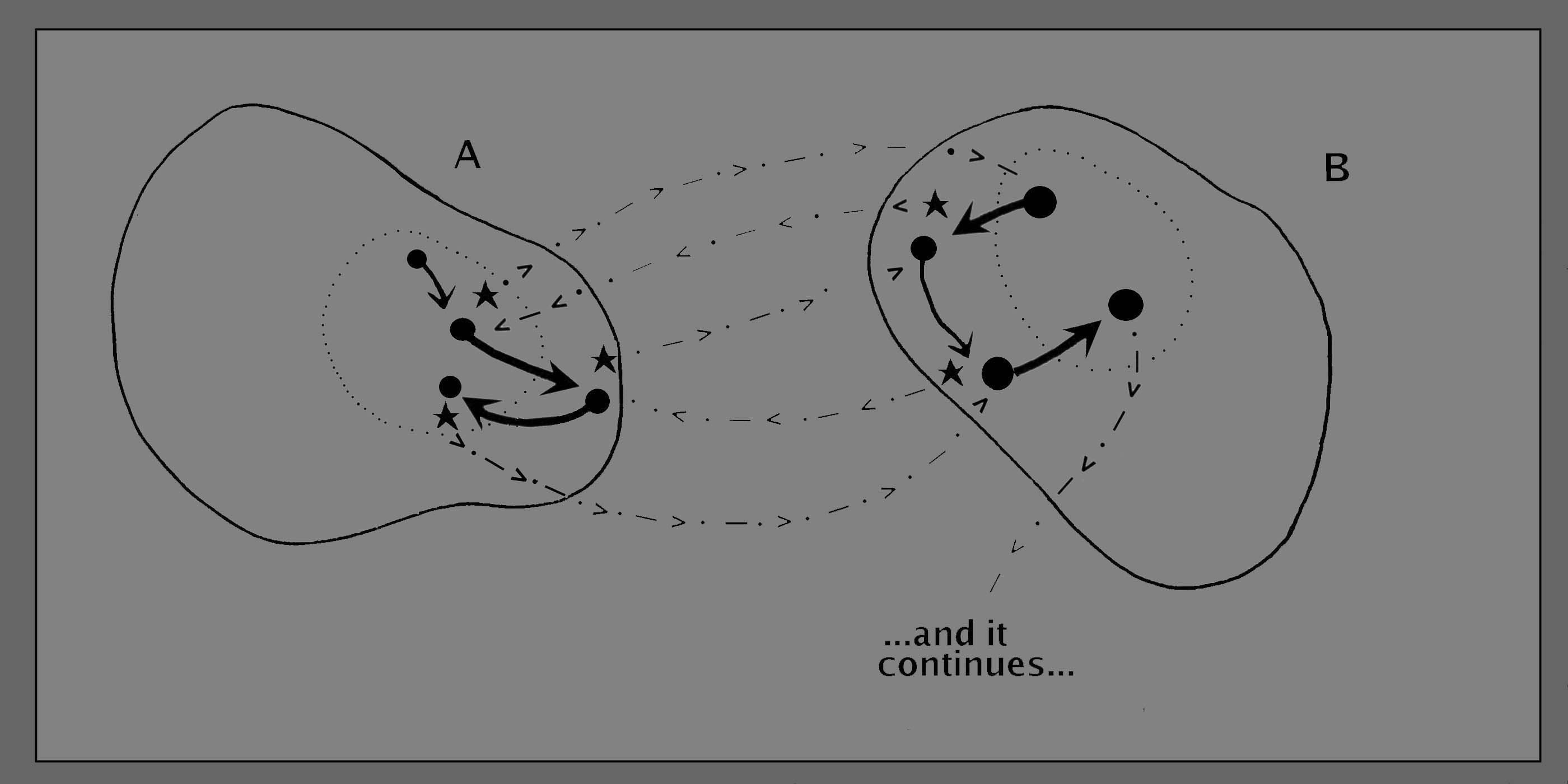
Figure
7
So what we
are
looking at is a process of structural coupling between two
organisms which follows automatically from their arrival in each
other's neighbourhood. Maturana and Varela point out that this is a
new level of complexity in its own right, and gives rise to what they
call a consensual domain. The patterns of behaviour in this
domain amount to a private language having developed spontaneously
between the two participants, and it is therefore also termed a domain
of linguistic behaviour. This is really an extraordinary
situation for us to contemplate, because most current theories of
language assume that there has to be a pre-established code or
convention before people can communicate through language. Yet here we
have two very simple-minded creatures who seem to be capable of
entering into their own linguistic community, establishing whatever
codes or conventions they need as they go along!
With respect
to our
own exploration of facts, feelings and their inter-relationships we
need to be clear that these unicellular organisms are communicating
exclusively in terms of feelings at the stage I have
described. Each message is, in effect: "This is what it feels like for
me, to be on the receiving end of what you just did!" However, as
Maturana and Varela point out, an exclusively connotational language
such as this can still give rise to a subtle and precise mutual
orientation. For instance we need very little further elaboration of
the theory to provide a satisfactory account of the honey bees' dance -
a dance which can orient a swarm of bees to a particular patch of a
particular species of flower from several miles away(3).
Our only problem is that in order to read the dance we would
have to have the inner neurological structure of a honey bee - in which
case we might find it very difficult to communicate our findings to our
human colleagues!
What we have
found
here is something resembling a spontaneous intelligence, an automatic
process of coming-into-tune, which works at the level of the simplest
organic processes. It does not require any consciousness, nor
intelligence as we normally understand the word, in order to operate.
Our working assumption in the chapters which follow is going to be that
this rudimentary kind of intelligence is working busily within us at
every moment of our lives, and that it underpins our higher-level
faculties in ways which we normally take for granted. The implications
of this assumption are extensive, but there are two major challenges
entailed in it which I want to take special note of.
Firstly, we
need to
learn to recognize when and where this organic intelligence can be
trusted to do its work, free from unnecessary interference. This is so
that the higher levels of our intentional behaviour are enabled to work
in harmony with the lower levels, in contrast with that sad but common
situation where we frustrate our own best intentions by pushing against the
deeper flow.
The other
challenge
is to identify those sensibilities, empathies and rationalities which
are most distinctly human, and try to better understand the
relationship between these and the organic intelligence which underlies
them.
Please Note:-
this section begins a sequence of sustained argument, linking the
theory of auto-poiesis to the radical personal and cultural enquiry
which is the sphere of interest of this web-site overall. The argument
continues under the following headings:-
NOTES
TO
THIS SECTION
1. ROTH, Michael. A Bolt From the Bleeding Sky,
Dielectric Publications, London 1984
2. The
importance of "tacit knowledge" in the performance of every
human skill is one of the central arguments in Michael Polanyi's
authoritative and influential book Human Knowledge. Tacit
knowing is what apprentices learn on their long years of training, and
something the master craftsman can rarely put into words. This
non-conscious level of "knowing what to do" is the exact counterpart of
Maturana and Varela's structural coupling.
3. .I
would hypothesise that the structural coupling which enables this
highly specialised and effective communication has been built up over
many billions of years of single-minded practice. The precise nature
of the coupling can only be traced by means of painstaking observation
and hypothesis, of course.
© all content: copyright reserved,
Michael Roth, March 2009







